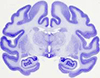
The work deals with cytological characteristics of neurons, glias and synapses of the anterior amygdaloid area of rat brain amygdaloid complex, one of the areas of sexual dimorphism, at the stage of estrous and after gonadectomy. 

In the forebrain, the highest density of hFF2 expressing neurons was observed in the anterior amygdaloid area and dorsomedial hypothalamic nucleus, especially in its caudal part.
In the amygdaloid complex, GAD65 is strongly expressed in striatal-like divisions, namely, the anterior amygdaloid area, the central nucleus (CEA), the intercalated nuclei, and the dorsal part of the medial nucleus (MEA).
The anterior amygdaloid area contained numerous NADPHd-positive neurons; in its dorsal part the majority of cells were only moderately stained, whereas in the ventral part the neurons were very strongly stained.
Cholinergic basal forebrain neurons confined within the intrinsic connections of the extended amygdala in the caudal sublenticular region and anterior amygdaloid area (cSLR/AAA) differ from other basal forebrain cholinergic neurons in several morphological and neurochemical respects.
(3) The anterior amygdaloid area and the corticomedial and basolateral complexes appear at stage 16.
We report that the afferent projections to the subparafascicular nucleus and area include the medial prefrontal, insular, and ectorhinal cortex, the subiculum, the lateral septum, the anterior amygdaloid area, the medial amygdaloid nucleus, the caudal paralaminar area of the thalamus, the lateral preoptic area, the anterior, ventromedial, and posterior hypothalamic nuclei, the dorsal premamillary nucleus, the zona incerta and Forel's fields, the periaqueductal gray, the deep layers of the superior colliculus, cortical layers of the inferior colliculus, the cuneiform nucleus, the medial paralemniscal nucleus, and the parabrachial nuclei.
The highest density of 5-hydroxytryptamine immunoreactive fibers is observed in the central nucleus, the nucleus of the lateral olfactory tract, the paralaminar nucleus, the anterior amygdaloid area and a small region of the amygdalohippocampal area.
The entire amygdaloid complex was outlined and then further partitioned into five reliably defined subdivisions: 1) the lateral nucleus, 2) the basal nucleus, 3) the accessory basal nucleus, 4) the central nucleus, and 5) the remaining nuclei (including anterior cortical, anterior amygdaloid area, periamygdaloid cortex, medial, posterior cortical, nucleus of the lateral olfactory tract, amygdalohippocampal area, and intercalated nuclei).
We first report CGRP terminal fields in the olfactory-anterior septal region and also CGRP projections from the parabrachial nuclei to the olfactory-anterior septal region, the medial prefrontal cortex, the interstitial nucleus of the anterior commissure, the nucleus of the lateral olfactory tract, the anterior amygdaloid area, the posterolateral cortical amygdaloid nucleus, and the dorsolateral part of the lateral amygdaloid nucleus.
A lesser density was revealed in the dorsomedial hypothalamic nucleus, basal nucleus of Meynert, and anterior amygdaloid area.
Anisotropic flow along the external capsule tracts carried GDNF into the anterior amygdaloid area.
In addition PSA-NCAM immunoreactive neuronal somata and processes exist in the lateral, basal and accessory basal nuclei, anterior amygdaloid area and amygdalo-striatal area.
Reactivity for phosphorylase a, the active form of glycogen phosphorylase, was higher in all parts of the medial amygdaloid nucleus, in the medial division of the central amygdaloid nucleus, in the anterior amygdaloid area and in the bed nucleus of stria terminalis than in all parts of the lateral amygdaloid nucleus, the anterior cortical amygdaloid nucleus, the posteromedial and posterolateral cortical amygdaloid nuclei, the intercalated nucleus of the amygdala, main part and the intercalated nuclei.
In 3-Br-7NI-treated relative to oil-treated females, the number of citrulline-positive cells was reduced by 70% in the PVN and by 50% in the anterior amygdaloid area, a control region of the brain.
Retrogradely labeled cells are found in insular, infralimbic, prelimbic, piriform, amygdalopiriform transition, entorhinal and perirhinal cortices, as well as in temporal field CA1 of Ammon horn and ventral subiculum, amygdala (nucleus of the lateral olfactory tract, anterior amygdaloid area, anterior cortical, posterolateral cortical, anterior and posterior basomedial, intercalated cells, basolateral and lateral nuclei), and extended amygdala, primarily in its central division.
After infusions of SeO3(2-) into combinations of cortical, medial, or amygdalohippocampal regions, retrogradely labeled zinc-containing somata were found in all amygdaloid nuclei except for the medial and central nuclei, the bed nucleus of the accessory olfactory tract, the nucleus of the lateral olfactory tract, and the anterior amygdaloid area.
Some corticotropin-releasing hormone-labeled cells are scattered in other subdivisions of the central nucleus and throughout the anterior amygdaloid area.
Light projections were observed in the parvicellular division of the basal nucleus, the anterior cortical nucleus, the amygdalohippocampal area, and the anterior amygdaloid area.
Sparse alpha-melanocyte-stimulating hormone-immunoreactive fibers were found in the basomedial nucleus of the amygdala, whereas a low density of fibers containing alpha-neo-endorphin was observed in the anterior amygdaloid area. Neurotensin was observed in fibers of the anterior amygdaloid area (low density) and both the lateral (low density) and the medial part (moderate density) of the central nucleus. A low density of fibers containing neurokinin A was found in the anterior amygdaloid area, the basolateral nucleus, and the medial part of the central nucleus.
Compared with previously reported distributions, our reinvestigation describes novel oxytocin- and vasopressin-binding sites in the lateral and supracapsular BST, in the sublenticular extended amygdala, in the interstitial nucleus of the posterior limb of the anterior commissure, in the marginal zone, in the central amygdaloid nucleus, and in the anterior amygdaloid area.
The parvicellular division projected lightly to the homonymous region on the contralateral side, to the contralateral anterior amygdaloid area and to the medial division of the central nucleus.
The present results confirm previously reported afferent connections of the infralimbic area and further reveal additional inputs from the anterior amygdaloid area, anterior cortical amygdaloid nucleus, posterolateral cortical amygdaloid nucleus, ventral reunions, lateral preoptic area, dorsal hypothalamic area, posterior hypothalamic nucleus, supramammillary nucleus and medial mammillary nucleus.
High densities of 125I-NT binding sites were found in the following amygdaloid structures the dorsal part of the accessory basal nucleus, the medial part of the cortical nucleus, the lateral subdivision of the central nucleus, the paralaminar nucleus, the amygdalohippocampal transition area and the rostral portions of the anterior amygdaloid area. The ventral part of the accessory basal nucleus, the intercalated cell groups and the remaining parts of the anterior amygdaloid area showed moderate density of NT binding sites, while the medial, basal and lateral amygdaloid nuclei, the lateral part of the cortical nucleus, the medial subdivision of the central nucleus, as well as the corticoamygdaloid transition area exhibited low densities of 125I-NT binding sites.
Injections into the infralimbic cortex produced anterograde labeling in the lateral capsular subdivision of the central nucleus, superficial (corticomedial) amygdaloid nuclei, lateral and accessory basal nuclei, and the anterior amygdaloid area.
The main intraamygdaloid targets of the basal nucleus projections are the nucleus of the lateral olfactory tract, the anterior amygdaloid area, the medial and capsular divisions of the central nucleus, the anterior cortical nucleus, and the amygdalohippocampal area.
Very few labeled cell bodies were seen in the anterior amygdaloid area and the medial nucleus.
The density of choline acetyltransferase-positive fibers was high in the nucleus of the lateral olfactory tract, the basolateral nucleus, and the amygdalohippocampal area; medium in the lateral nucleus, the cortical nucleus, the accessory basal nucleus, the periamygdaloid cortex, and the anterior amygdaloid area; and low in the medial and central nuclei. Nerve growth factor receptor-positive fibers were of medium density in the lateral nucleus, the accessory basal nucleus, the cortical nucleus, the anterior amygdaloid area, the periamygdaloid cortex, and the amygdalohippocampal area.
In the present studies, each of the melanocortin peptides alpha-MSH, des-acetyl-alpha-MSH, beta-MSH and ACTH1-24 when present at 1 microM virtually eliminated [ 125I]NDP-MSH binding in each of a series of brain structures, including medial preoptic area, caudate putamen, olfactory tubercle, bed nucleus of the stria terminalis, ventral part of the lateral septal nucleus, hypothalamic periventricular and paraventricular nuclei, dorsal anterior amygdaloid area, substantia innominata and thalamic paraventricular nucleus; as well as in extraorbital lacrimal gland, a peripheral melanocortin target.
The basomedial, accessory basal, and cortical nuclei, the amygdalohippocampal and cortico-amygdaloid transition areas, as well as the anterior amygdaloid area, showed a moderate density of ChAT-positive varicosities and fibers.
After a 750-V shock it was the substantia nigra, the lateral septal nucleus, the anterior amygdaloid area, and the lateral preoptic area that consistently contained electrically damaged neurons, without other kinds of parenchymal damage..
The distribution and neuronal morphology of labeled neurons in the central nucleus and anterior amygdaloid area suggests that diaphorase-positive cells in these areas may be cholinergic.
Aromatase activity was measured separately in homogenates of left and right preoptic area, anterior amygdaloid area, medial amygdaloid nucleus, anterior hypothalamic area and posterior hypothalamic area, by the tritiated water method with [ 1 beta-3H]-androstenedione as a substrate.
The anterior amygdaloid area, basal complex, paralaminar nucleus, cortical nucleus, cortical-amygdaloid transition area, and amygdalohippocampal area contained moderate densities of immunoreactivity. Immunoreactive neurons were found only within the anterior amygdaloid area and the central, medial, intercalated, and lateral capsular nuclei.
The magnocellular division of the medial geniculate, a thalamic auditory (and, to a moderate degree, a spinothalamic) relay, sent heavy projections to the central, accessory basal, lateral, and anterior cortical nuclei, and to the anterior amygdaloid area and the nucleus of the accessory olfactory tract.
The highest densities of stained fibers were found in the lateral nucleus, the parvicellular portion of the accessory basal nucleus, and the anterior amygdaloid area.
The complex of the MFB-related areas is surrounded by the following cellular entities: the nucleus accumbens (ACB), the caudatus-putamen region (CPU), the globus pallidus (GP), the bed nucleus of the stria terminalis (BST), the anterior amygdaloid area (AAA), the amygdaloid nuclear complex (A), the medial preoptic area (MPOA) and the anterior hypothalamic area (AHA).
Immunocytochemically, chromogranin A was also detected in the central projection areas of mitral and tufted cells in the primary olfactory cortex and the anterior amygdaloid area but not in the olfactory glomeruli, where the incoming olfactory nerve fibers of the primary olfactory neurons establish synaptic contacts.
There was also variation of staining intensity within many of the nuclear subdivisions, such as the lateral and central nuclei, anterior amygdaloid area and the intercalated groups.
In pregnant females the anterior amygdaloid area and the bed nucleus of the stria terminalis were labelled additionally. After the first 5 days of lactation, estrogen-binding cells could no longer be detected in the hippocampus, entorhinal and piriform cortex and anterior amygdaloid area.
After different dye injections in both MOBs, labeled cells were present in the following structures: tenia tecta, vertical limb of the diagonal band of Broca, and medial septal nucleus in the ipsilateral injection site; and the NOA, piriform cortex, nucleus of the lateral olfactory tract, horizontal limb of the diagonal band of Broca, posterolateral cortical amygdaloid nucleus, anterior amygdaloid area, and dorsal raphe nucleus in both hemispheres. Structures showing double-labeled cells were the NOA, horizontal limb of the diagonal band of Broca, nucleus of the lateral olfactory tract, anterior amygdaloid area, and posterolateral cortical amygdaloid nucleus. After HRP injections in the dorsal part of the NOA, labeled cells were distributed in the NOA, nucleus of the lateral olfactory tract, posterolateral cortical amygdaloid nucleus, piriform cortex, horizontal and vertical limbs of the diagonal band of Broca, mitral cell layer of the MOB, tenia tecta, anterior amygdaloid area, and the contralateral NOA.
Our study revealed a characteristic distribution of GABAergic and cholinergic elements in the various amygdaloid nuclei: 1) Large, ChAT-immunopositive cells with heavily labeled dendrites were observed in the anterior amygdaloid area and in the lateral and medial zones of the central nucleus.
As defined by these markers, the ventral pallidum was traced rostroventrally from the globus pallidus to the superficial layers of the olfactory tubercle, medially to the insula Calleja magna and the lateral septum, laterally to the pyriform cortex, and caudally to the anterior amygdaloid area.
The ventral SI projects more heavily to the medial part of the bed nucleus of the stria terminalis; the anterior amygdaloid area; a ventromedial amygdaloid region that includes but is not limited to the medial nucleus; the lateral and medial preoptic areas; and the anterior hypothalamus.
In addition to the hypothalamic projections, there is in the hamster a small projection along the base of the telencephalon to the anterior amygdaloid area and cortical amygdaloid nucleus and a very sparse projection to the anterior thalamic nuclei..
In contrast to the caudate nucleus, nucleus accumbens, and olfactory tubercle, neither gammabutyrolactone (GBL) nor axotomy increased biochemically estimated DA synthesis (DOPA accumulation) in any of the amygdaloid nuclei, the anterior amygdaloid area, septal nuclei, or subdivisions of the interstitial (bed) nucleus of the stria terminalis.
Additional retrogradely labeled cells were found in the endopiriform nucleus, the anterior amygdaloid area, and the cortical nuclei.
Additional cells were seen in the central nucleus, basomedial nucleus and anterior amygdaloid area.
By means of cobalt-enhanced immunohistochemistry, CRFI cells were detected in almost all the regions of the amygdala, including the central amygdaloid nucleus (Ce), basolateral amygdaloid nucleus (B1), intra-amygdaloid bed nucleus of the stria terminalis (Abst), medial amygdaloid nucleus (Me), amygdalohippocampal area (Ahi), posterior cortical amygdaloid nucleus (Aco), lateral amygdaloid nucleus (La), anterior amygdaloid area (AAA) and basomedial amygdaloid nucleus (Bm).
Each of the basal nuclei project ipsilaterally to the anterior amygdaloid area, substantia innominata and topographically to the ventral part of the striatum and adjacent olfactory tubercle.
The infralimbic area does not project to the basolateral nucleus and cortico-amygdaloid projections from this area are focussed on the anterior cortical nucleus and the anterior amygdaloid area. Based on similar projections, this area appears to be a caudal continuation of the anterior amygdaloid area.
injection of [ 3H]estradiol revealed labeled cells in all of the above regions, in addition to the basolateral portion of the septum, the bed nucleus of the stria terminalis, and in the anterior amygdaloid area as well as the medial and cortical nuclei of the amygdala.
A few GAD-positive bulbopetal cells, furthermore, are located in the ventral pallidum, anterior amygdaloid area, deep olfactory cortex, nucleus of the lateral olfactory tract, lateral hypothalamic area, and tuberomamillary nucleus.
Unilateral implants containing oestradiol benzoate (OB) and cholesterol at the ratio of 1 + 4 were placed for 48 h either in the region of the anterior amygdaloid area (AAA) and nucleus of the lateral olfactory tract (NLOT) or in the anterior or posterior part of the mediocortical amygdala of immature 26-day-old female rats, and the age at vaginal opening and the first ovulation was recorded.
Other olfactory centers of the hemisphere (anterior olfactory nucleus, olfactory tubercle, bandaletta diagonalis (Broca) nucleus and medial nucleus of the anterior amygdaloid area) send to the hippocamp their afferent fibres.
Interference between seizure activity and orthodromic focal potentials caused by stimulation of the periamygdaloid cortex and anterior amygdaloid area was shown.
Injections confined to the ventromedial nucleus also result in labeling of the piriform cortex, the periamygdaloid cortex, the anterior amygdaloid area, the medial amygdaloid nucleus and the parvocellular divisions of both the basal and basal accessory nuclei. Isotope injections that involve the caudal part of the lateral hypothalamic area label projections to the medial division of the central amygdaloid nucleus, to the medial and cortical nuclei and to the anterior amygdaloid area.
As demonstrates the analysis of the data obtained, in male rats the following area respond to gonadectomy: neurons of the anterior amygdaloid area of the dep zone of the anterior cortical nucleus, of the central nucleus, of the posterior intercalated masses, of the dorsomedial nucleus, of the posterior medial nucleus and of the medial part in the posterior cortical nucleus.
NT-positive structures first appear in the primordium of the olfactory bulb, anterior amygdaloid area, piriform cortex, amygdaloid complex, nucleus tractus diagonalis, lateral preoptic area, and lateral hypothalamus, etc., of the fetus of gestational day 16.
Terminal degeneration has also been observed in the deep layers of the anterior olfactory nucleus, the olfactory tubercle, the nucleus of the lateral olfactory tract, and the anterior amygdaloid area.
Fibers originating in the medial part of the parabrachial nucleus in addition have a dense termination in the anterior amygdaloid area.
The anterior amygdaloid area shows a similar projection field, the only difference being that this structure does not project to any parts of the medial nucleus.
After injections which involve Ammon's horn, the dentate gyrus, and the subicular complex, retrogradely labeled neurons are found in the following regions: in the amygdala (specifically in the anterior amygdaloid area, the basolateral nucleus, and the periamygdaloid cortex); in the medial septal nucleus and the nucleus of the diagonal band; in the ventral part of the claustrum; in the substantia innominata and the basal nucleus of Meynert; in the rostral thalamus (specifically in the anterior nuclear complex, the laterodorsal nucleus, the paraventricular and parataenial nuclei, the nucleus reuniens, and the nucleus centralis medialis); in the lateral preoptic and lateral hypothalamic areas, and especially in the supramammillary and retromammillary regions; in the ventral tegmental area, the tegmental reticular fields, the raphe nuclei (specifically in nucleus centralis superior and the dorsal raphe nucleus), in the nucleus reticularis tegmenti pontis, the central gray, the dorsal tegmental nucleus, and in the locus coeruleus..
Cells labeled with the peroxidase reaction product were located in gyrus propreus, gyrus genualis, nucleus accumbens, bed nucleus of the stria terminalis, bed nucleus of the anterior commissure, nucleus of the diagonal band, substantia innominata, anterior amygdaloid area, ventromedial hypothalamic area, paraventricular nucleus, perifornical hypothalamic area, lateral hypothalamic area, dorsal hypothalamic area, field of Forel, midbrain reticular formation, superior colliculus, ventral central grey, lateral central gray, locus coeruleus, parabrachial nuclei, nucleus of the lateral lemniscus, oral pontine reticular nucleus, and the dorsal raphe.
Labelled fibers and terminal fields were seen in the preoptic area, bed nucleus of the stria terminalis, substantia innominata, the anterior amygdaloid area, diagonal bands of Broca and lateral septum.
A moderate to small number of labeled cells, furthermore, were seen in the dorsal (DT) and medial (MT) transition fields, the ventral praecommissural hippocampus (tt2), the ventral superficial part of the nucleus of the vertical limb of the diagonal band (NVDB), the sublenticular part of the substantia innominata (SI), the anterior amygdaloid area, the posterolateral cortical amygdaloid nucleus (C2) and the transition region (28 L') between the olfactory cortex and the lateral entorhinal area proper.
The large NE fibers appear to enter the amygdala via the ansa peduncularis-ventral amygdaloid bundle to innervate the central and basolateral nucleus and the anterior amygdaloid area.
The lateral, basal dorsal and basal ventral nuclei belong to the first, the medial and central nucleus and the anterior amygdaloid area--to the second group.
Injections throughout the prefrontal cortex labeled neurons in the ipsilateral basal magnocellular amygdaloid nucleus; injections in the premotor and motor cortices labeled neurons in the same nucleus plus a few cells in the anterior amygdaloid area..
Other fibers coursed over the genu of the corpus callosum, through the stria terminalis and ansa peduncularis to the medial, cortical and basal amygdaloid nuclei and the anterior amygdaloid area, and through the medial forebrain bundle to the substantia nigra.
There were no important changes in the vaginal patterns or ovarian and uterine morphology in animals given injections in the same area with the solvent only and in animals given injections with 6-OHDA in the anterior amygdaloid area.
The ventral portion extended into the olfactory tuberculum and the anterior amygdaloid area.
Damage to the anterior amygdaloid area or the the nuclei of the lateral olfactory tract did not reliably affect active or passive avoidance behavior..
-
[ View All ]